Comment la production de calcite intracellulaire chez les algues de type coccolithophoridé est-elle approvisionnée en calcium ?
Pour former les coques écailleuses dures qui entourent leurs cellules, les algues de type coccolithophoridé produisent des écailles calcitiques dans un compartiment de calcification intracellulaire et les secrètent à la surface de la cellule. La formation des écailles nécessite le transfert de grandes quantités de calcium depuis l'eau de mer environnante jusqu'au compartiment de calcification. Les voies qui approvisionnent ce compartiment en calcium ne sont pas encore déterminées.
Pour la première fois, une équipe de chercheurs de l'Institut Max Planck de physiologie végétale moléculaire (Allemagne), de l'Institut Max Planck des colloïdes et des interfaces (Allemagne), de l'Institut allemand de recherche pour les géosciences (Allemagne) et de la ligne de lumière LUCIA ont mis en évidence qu’un compartiment récemment découvert, contenant une phase de phosphate de calcium (riche en Ca-P), participait à l'approvisionnement du compartiment de calcification en calcium.
Les coccolithophores sont des algues monocellulaires vivant dans les couches ensoleillées des océans du monde entier. Ces algues se caractérisent par des coques extracellulaires formées d'une multitude de petits disques minéralisés nommés coccolithes (Fig. 1A). Chaque coccolithe est constitué de cristaux nanoscopiques de calcite, un minéral produit à l'intérieur de la cellule au sein d'un compartiment spécialisé appelé la vésicule coccolithique. La plupart des aspects de la formation des coccolithes restent non élucidés, notamment les voies par lesquelles les ions constitutifs se déplacent pour finalement former la calcite coccolithique.
La formation biologiquement contrôlée de la calcite est un phénomène largement répandu et bien étudié chez les organismes multicellulaires. De récentes études sur les mécanismes de biominéralisation chez divers calcificateurs multicellulaires ont révélé une utilisation largement répandue de de phases intermédiaires amorphes dans la formation de biomatériaux cristallins. Dans le coccolithophore Emiliania huxleyi, l’organisme calcificateur le plus abondant sur Terre, l'équipe de chercheurs a très récemment découvert un compartiment stockant une phase amorphe riche en Ca-P. Cette découverte a soulevé la question de savoir si la réserve de Ca-P est une source de calcium pour la formation des coccolithes.
Pour traiter la question du transfert de matière entre le compartiment riche en Ca-P et la vésicule coccolithique, les chercheurs ont mené des expériences d'analyse pulse-chase avec du Sr (Fig. 1B) et ont utilisé la spectroscopie d'absorption de rayons X (XANES) en conditions cryogéniques, ainsi que la microscopie électronique en transmission pour suivre le Sr à l'intérieur des cellules de E. huxleyi. Les ions Sr ont servi de marqueur du Ca car ils sont transportés par des canaux de Ca et peuvent être transférés dans la vésicule coccolithique, comme le montre l'incorporation du Sr dans la calcite coccolithique.
L'étude de coupes minces de E. huxleyi par microscopie électronique a mis en évidence le fait que le Sr s'accumule dans la calcite coccolithique ainsi que dans le compartiment riche en Ca-P, mais pas dans d'autres emplacements intracellulaires. Ceci montre qu’en dehors de la vésicule coccolithique, le compartiment riche en Ca-P renferme la majorité du Sr intracellulaire.
Les chercheurs ont utilisé la spectroscopie XANES au seuil L2,3 du Sr en conditions cryogéniques pour déterminer les environnements chimiques du Sr cellulaire à différentes étapes des expériences d'analyse pulse-chase (Fig. 1C). En parallèle à l'impulsion Sr, la formation coccolithique a été commutée entre les états inactif et actif. Les résultats des expériences de pulse-chase ont montré que l'environnement du Sr dans les cellules non calcificatrices (blocage de la formation des coccolithes) est très semblable à celui du Sr en phase amorphe. À mesure que la calcification commence dans la période de chasse, le signal du Sr devient semblable à celui du Sr incorporé dans la calcite (Fig. 1D,E).
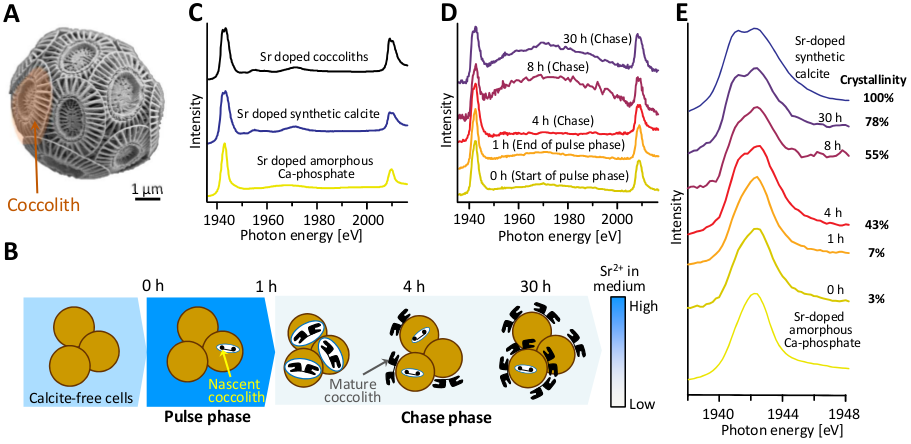
Ayant constaté que les ions Sr sont initialement présents dans l'environnement amorphe de la phase riche en Ca-P et passent à un environnement cristallin à mesure que la calcification progresse, les conclusions des expériences sont que le Ca est transféré depuis la réserve riche en Ca vers la vésicule coccolithique, et d'autre part que le compartiment riche en Ca-P entre dans la voie de biogénèse des coccolithes.
Ces résultats donnent non seulement un éclairage fondamental sur les mécanismes de la formation des coccolithes, mais ont aussi des conséquences au-delà du domaine de la biominéralisation.
Cette nouvelle perspective sur l'approvisionnement en matière de la vésicule coccolithique est en effet pertinente pour l'interprétation de la teneur en métaux trace des coccolithes fossiles dans le contexte des reconstructions paléoclimatiques, ainsi que pour la compréhension et la prédiction de la réponse des coccolithophores aux changements environnementaux à l'échelle mondiale, tels que l'acidification des océans.